Endosimbiosis seriada
]La teoría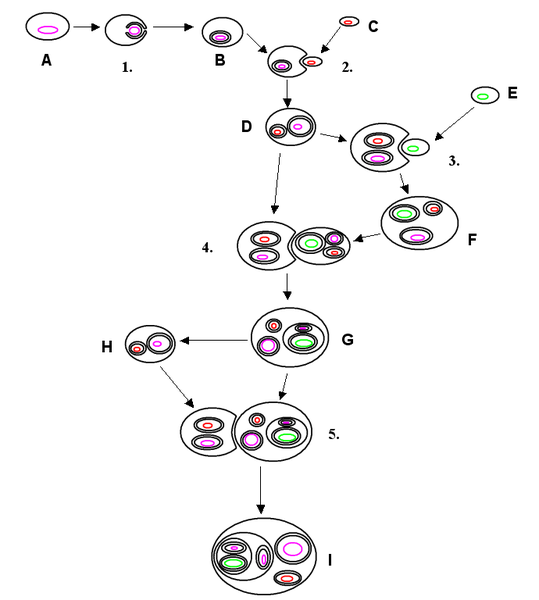
La teoría endosimbiótica describe el paso de las células
procariotas(bacterias o arqueas, no nucleadas) a las células
eucariotas (células nucleadas constituyentes de todos los pluricelulares)
mediante incorporaciones simbiogenéticas.
Margulis describe este paso en una serie de tres incorporaciones
mediante las cuales, por la unión simbiogenética de bacterias, se originaron
las células que conforman a los individuos de los otros cuatro reinos (protistas, animales, hongos y plantas).
Según la estimación más aceptada, hace 2.000 millones de años
(aunque una horquilla posible podría descender a la cifra de 1.500 millones de
años) la vida la componían multitud de bacterias diferentes, adaptadas a los
diferentes medios. Margulis destacó también, la que debió ser una alta
capacidad de adaptación de estas bacterias al cambiante e inestable ambiente de
la Tierra en
aquella época. Hoy se conocen más de veinte metabolismos diferentes usados por
las bacterias frente a los dos utilizados por los pluricelulares: el aeróbico,
que usa el oxígeno como fuente de energía -único metabolismo utilizado por los
animales- y la fotosíntesis -presente en las plantas-. Para Margulis, tal
variedad revela las dificultades a las que las bacterias se tuvieron que
enfrentar y su capacidad para aportar soluciones a esas dificultades.
A mediados
de los sesenta, Margulis formuló lo que se conoce como «Teoría de la
endosimbiosis serial», que propone que la primera célula eucariota de la
Tierra, aquella célula de la que provenimos todos los animales y las plantas,
se formó mediante la fusión de tres bacterias preexistentes completas, con los
genes de cada una incluidos, por supuesto. Una de esas bacterias aportó los
andamios de microtúbulos, otra ciertas capacidades metabólicas peculiares y la
tercera (que se sumó más tarde a las otras dos) se convirtió en las actuales
mitocondrias. Esa célula eucariota primigenia empezó a proliferar, y una de sus
descendientes sufrió aún otra experiencia traumática: se tragó a una bacteria
fotosintética de la que provienen los actuales cloroplastos.
Javier Sampedro, Deconstruyendo
a Darwin.14
Primera incorporación simbiogenética:
En primer
lugar, un tipo de bacteria amante del azufre y del calor, llamada
arqueobacteria fermentadora (o termoacidófila), se fusionó con una bacteria
nadadora. Juntos, los dos componentes integrados de la fusión se convirtieron
en el nucleocitoplasma, la sustancia base de los ancestros de las células
animales, vegetales y fúngicas. Este temprano protista nadador era, como sus
descendientes actuales, un organismo anaerobio. Envenenado por el oxígeno,
vivía en arenas y lodos donde abundaba la materia orgánica, en grietas de las
rocas, en charcos y estanques donde este elemento estaba ausente o era escaso.
Una bacteria consumidora
de azufre, que utilizaba el azufre y el calor como
fuente de energía (arquea fermentadora o termoacidófila), se
habría fusionado con una bacteria nadadora (espiroqueta) habiendo
pasado a formar un nuevo organismo y sumaría sus características iniciales de
forma sinérgica (en la que el resultado de la incorporación de dos o más
unidades adquiere mayor valor que la suma de sus componentes). El resultado
sería el primer eucarionte (unicelular eucariota) y ancestro
único de todos los pluricelulares. El núcleoplasma de la células de animales,
plantas y hongos sería el resultado de la unión de estas dos bacterias.
A las características
iniciales de ambas células se le sumaría una nueva morfología más compleja con una nueva y llamativa
resistencia al intercambio genético horizontal. El ADN quedaría
confinado en un núcleo interno separado del resto de la célula por
una membrana.
Sobre este primer paso, al
día de hoy, todavía existen discrepancias. A finales de los años ochenta y
principio de los noventa diversos trabajos no admitían las homologías
propuestas entre los flagelos de los eucariontes y de las espiroquetas.
Segunda incorporación
simbiogenética:
Después de que evolucionara la mitosis en los protistas nadadores,
otro tipo de microorganismo de vida libre fue incorporado a la fusión: una
bacteria que respiraba oxígeno. Surgieron células todavía más grandes, más
complejas.
Este nuevo organismo
todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este
gas suponía un veneno para
él, por lo que viviría en medios donde este oxigeno, cada vez más presente,
fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio
eucarionte de la capacidad para metabolizar oxigeno. Este nuevo endosombionte,
originariamente bacteria respiradora de oxigeno de vida libre, se convertiría
en las actuales mitocondrias y peroxisomas presentes en las células eucariotas de
los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha
llegado a convertirse el planeta Tierra. Los animales y hongos somos el
resultado de esta segunda incorporación.
Tercera incorporación simbiogenética:
En la adquisición final de la serie generadora de células
complejas, los respiradores de oxígeno engulleron, ingirieron, pero no pudieron
digerir bacterias fotosintéticas de color verde brillante. La «incorporación»
literal tuvo lugar tras una gran lucha en la que las bacterias verdes no
digeridas sobrevivieron y la fusión completa prevaleció. Con el tiempo las
bacterias verdes se convirtieron en cloroplastos (paso 4, figura 1.1). Como
cuarto miembro, estos productivos amantes del sol se integraron con los demás
socios anteriormente independientes. Esta fusión final dio lugar a las algas
verdes nadadoras. Estas antiguas algas verdes nadadoras no sólo son los
ancestros de las células vegetales actuales; todos sus componentes individuales
todavía están vivos y en buena forma, nadando, fermentando y respirando
oxígeno.
Margulis, Una
revolución en la Evolución, Cap.: Individualidad
por incorporación.
Esta tercera incorporación
originó el Reino vegetal, las recientemente adquiridas células respiradoras de
oxígeno fagocitarían bacterias fotosintéticas y algunas de ellas, haciéndose
resistentes, pasarían a formar parte del organismo, originando a su vez un
nuevo organismo capaz de sintetizar la energía procedente del Sol. Estos nuevos pluricelulares, las plantas,
con su éxito, contribuyeron y contribuyen al éxito de animales y hongos.
En la actualidad permanecen las bacterias descendientes de
aquellas que debieron, por incorporación, originar las células eucariotas; así
como aquellos protistas que no participaron en alguna de las sucesivas
incorporaciones.
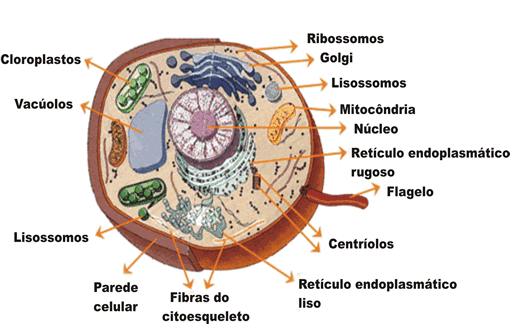
La evidencia de que las mitocondrias y los plastos surgieron a
través del proceso de endosimbiosis son las siguientes:
§ El tamaño
de las mitocondrias es similar al tamaño de algunas bacterias.
§ Las
mitocondria y los cloroplastos contienen ADN bicatenario
circular cerrado covalentemente - al igual que los procariotas- mientras que el
núcleo eucariota posee varios cromosomas bicatenarios lineales.
§ Están
rodeados por una doble membrana, lo que concuerda con la idea de la
fagocitosis: la membrana interna sería la membrana plasmática originaria de la
bacteria, mientras que la membrana externa correspondería a aquella porción que
la habría englobado en una vesícula.
§ Las
mitocondrias y los cloroplastos se dividen por fisión binaria al igual que los procariotas (los
eucariotas lo hacen por mitosis). En algunas algas, tales como Euglena, los plastos pueden ser destruidos por
ciertos productos químicos o la ausencia prolongada de luz sin que el resto de
la célula se vea afectada. En estos casos, los plastos no se regeneran.
§ En
mitocondrias y cloroplastos los centros de obtención de energía se sitúan en
las membranas, al igual que ocurre en las bacterias. Por otro lado, los
tilacoides que encontramos en cloroplastos son similares a unos sistemas
elaborados de endomembranas presentes en cianobacterias.
§ En
general, la síntesis proteica en mitocondrias y cloroplastos es autónoma.
§ Algunas
proteínas codificadas en el núcleo se transportan al orgánulo, y las
mitocondrias y cloroplastos tienen genomas pequeños en comparación con los de
las bacterias.. Esto es consistente con la idea de una dependencia creciente
hacia el anfitrión eucariótico después de la endosimbiosis. La mayoría de los
genes en los genomas de los orgánulos se han perdido o se han movido al núcleo.
Es por ello que transcurridos tantos años, hospedador y huésped no podrían
vivir por separado.
§ En
mitocondrias y cloroplastos encontramos ribosomas 70s, característicos de
procariotas, mientras que en el resto de la célula eucariota los ribosomas son
80s.
§ El
análisis del RNAr 16s de la subunidad pequeña del ribosoma de mitocondrias y
plastos revela escasas diferencias evolutivas con algunos procariotas.
§ Una
posible endosimbiosis secundaria (es decir, implicando plastos eucariotas) ha
sido observado por Okamoto e Inouye (2005). El protista heterótrofo Hatena se
comporta como un depredador e ingiere algas verdes, que
pierden sus flagelos y citoesqueleto, mientras que el protista, ahora un
anfitrión, adquiere nutrición fotosintética, fototaxia y pierde su aparato de
alimentación.
Las
bacterias, fusionadas en simbiosis, nos dejan pistas de su anterior
independencia. Tanto las mitocondrias como los plastos son bacterianos en su
tamaño y forma.
Margulis, Planeta
simbiótico.30


{kind=link}
{kind=link}